Tonny W Naranjo1,2 ![]() ,
Damaris Lopera2,
Andrés Felipe Zuluaga3,
Luz Elena Cano4
,
Damaris Lopera2,
Andrés Felipe Zuluaga3,
Luz Elena Cano4
For correspondence:- Tonny Naranjo Email: tonny.naranjo@upb.edu.co Tel:+5744035950
Received: 21 July 2016 Accepted: 17 November 2016 Published: 21 December 2016
Citation: Naranjo TW, Lopera D, Zuluaga AF, Cano LE. Immunomodulatory activity of itraconazole in lung. Trop J Pharm Res 2016; 15(12):2603-2609 doi: 10.4314/tjpr.v15i12.10
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To evaluate the in vivo effects of ITC as an immunomodulator on various immunological mediators, including cytokines, chemokines and growth factors in pulmonary homogenates.
Methods: Two experimental groups consisting of 25 BALB/c mice each were used: (a) PBS-treated mice and (b) ITC-treated mice (1 mg/mouse/day). The animals were treated daily via gavage for 8 weeks. Five mice from each group were sacrificed at 0, 4, 8, and 12 weeks and cytokine levels assessed in the supernatants of lung macerates using the Bio-Plex system. Five lungs from each experimental group, after 8 weeks of treatment, were used for histopathological analysis.
Results: Compared with the control group, ITC-treated mice showed significant changes in the pulmonary levels of 50 % of the molecules evaluated: IL-4, IL-10, IL-12p70, IL-13, TNF-α, eotaxin, MIP-1α, MIP-2, and LIF were significantly upregulated. In contrast, IL-1β, IL-15, IL-2, IL-1α, and VEGF were downregulated.
Conclusion: Although histopathological examination did not show changes in lung cell infiltrates, ITC exerted a marked immunomodulatory effect at the pulmonary level in healthy BALB/c mice.
Introduction
The marked increase in the incidence of fungal infections in recent decades, principally due to the growing number of patients with AIDS and organ transplants, has awakened interest not only in developing new antifungal drugs but also in evaluating the effects that current antifungals have on the host immune response [1-3]. It is important to ensure that these agents per se do not exert negative effects on the host immune system.
Itraconazole (ITC), a broad-spectrum azole antifungal agent, is used widely to prevent and treat some pulmonary fungal infections as well as other medically important fungi, causing superficial, subcutaneous, and systemic infections in humans [1]. The mechanism of action of ITC is based on inhibiting 14-α-sterol demethylase, a cytochrome P450 fungal enzyme essential for the biosynthesis of ergosterol, an important component of the fungal cell membrane. [4]. ITC is considered fungistatic, rather than fungicidal, and its action should be complemented by the host immune response in such a way that the infectious agent is eliminated [5].
During a fungal infection, the host immune response against the pathogen occurs at different levels, with increasing specificity over time; phagocytic cells, macrophages, dendritic cells, and neutrophils are responsible for innate immunity, representing the first line of defense against fungal pathogens. These cells produce and secrete a wide array of molecules, such as chemokines, growth factors, and pro- and anti-inflammatory cytokines. Expression of these molecules is an important step in the development of adaptive immunity, which depending on the profile expressed, may constitute a protective immune response [6,7].
In addition to their antimicrobial actions, certain drugs, particularly antibiotics, have direct modulating effects on the immune response, as shown by both in vitro and in vivo studies. Although several papers have described the in vitro effects of some antifungals on the activities of macrophages, monocytes, and neutrophils from animals and humans [2,3,7,8], little is known about the in vivo effects of antifungal drugs.
Louie et al [9] suggested that the efficacy of certain antifungal agents may be related to their ability to induce cytokine production. Fidan et al [10] showed that fluconazole, an antifungal drug used frequently to treat Candida infections, increased the levels of chemokines, such as CCL3 and CCL4, in the supernatants of human mononuclear cell cultures.
Because antifungal drugs are widely used in patients who are immunocompromised to some degree, it is important to determine the possible effects of these therapies on the immune system in vivo [4,11]. Given that little is currently known about the modulating properties of ITC, the aim of the present study was to evaluate the potential immunomodulatory activity of this antifungal drug on the pulmonary levels of 28 molecules with immunological activity in healthy mice.
Methods
Experimental animals
In total, 50 isogenic male BALB/c mice, 6 - 7 weeks old, weighing 18 - 20 g, were obtained from the breeding colony of the Corporación para Investigaciones Biológicas (CIB). Mice were bred and maintained in a micro-isolator system (Super Mouse 1800) at 22 °C under a 12/12 h light/dark cycle. The animals were provided sterile food and acidified water (pH 3.0 - 3.5) in sterilized bottles ad libitum and sterile bedding. All animal handling and maintenance procedures used were approved by the Ethics Committee of CIB (Act.092), and in accordance with the guidelines of NIH Laboratory Animal Care [12].
Drug, dosing and trial design
The animals were divided into two groups (PBS control vs. ITC treatment) of 25 mice each. The ITC was administered at 1 mg/mouse/day, the dose necessary to achieve serum levels of 1 μg/mL, as confirmed using a bioassay measuring blood levels of ITC. Lower levels of ITC have been associated with antifungal therapy failure [13]. Mice received 100 µL PBS or 100 µL ITC oral solution (Sporanox Janssen-Cilag S.A) daily for 8 weeks via gavage, and five members of each group were sacrificed at 0, 4, 8, and 12 weeks; the 12th week corresponded to a post-treatment follow-up interval of 4 weeks.
Sacrifice of animals and sample processing
At each of the time points evaluated, five animals from each group were euthanized by intraperitoneal injection of 1 mL 2.5 % sodium pentothal (Sandoz Laboratories, Kundl, Austria [14]. After the death of the mice, the lungs were removed and homogenized individually using a tissue grinder (Tissue Tearer model 985-370, Biospec Products) in 2 mL of solution containing a protease inhibitor cocktail (pepsin 0.1 µM, leupeptin 0.1 µM, phenylmethyl sulfonide fluoride 1 mM, N-tosyl-L-phenylalanine chloromethyl ketone 0.2 mM, and (α)-p-methyl L lysine chloromethyl ketone 0.1 mM (Sigma), plus ethylenediaminetetra-acetic acid 1 nM (Merck, Germany)). Homogenates were stored on ice and then centrifuged (3000 rpm, 15 min, 4 °C), and the supernatants were removed and stored at −70 °C until assayed [14].
To evaluate any potential cellular inflammatory responses, five lungs from both ITC-treated and PBS-treated mice after 8 weeks of treatment were perfused and fixed in 10 % formaldehyde−PBS and embedded in paraffin wax. Coronal 5 µm thick sections of the entire lung were stained with hematoxylin and eosin (H&E) for histopathological analysis.
Measurement of molecules with immunologi-cal activity
Supernatants of lung homogenates were thawed immediately prior to being used. The protein concentration was determined in each sample by the Bradford method (Bio-Rad Laboratories) following the manufacturer’s protocol and then normalized to 1 mg/mL. A magnetic bead-based multiplex assay was used to quantify the levels of multiple molecules in each normalized sample following the manufacturer’s instructions (Bio-Plex Pro-Mouse Assay, Bio-Rad Inc., CA, USA). The following molecules were measured: IL-1α, IL-1β, IL-2, IL-3, IL-4, IL-5, IL-6, IL-9, IL-10, IL 12(p40), IL-12(p70), IL-13, IL-15, IL-17, IFN-γ, TNF-α, eotaxin, granulocyte colony stimulating factor (G-CSF), granulocyte-monocyte colony stimulating factor (GM CSF), macrophage inflammatory proteins 1α, 1β, and 2 (MIP-1α, MIP-1β, MIP 2), fibroblast growth factor (FGF), vascular endothelial growth factor (VEGF), platelet-derived growth factor (PDGF), keratinocyte chemoattractant (KC), monokine induced by gamma interferon (MIG), and leukemia inhibitory factor (LIF).
Levels were determined using a multiplex array reader from Luminex Instrumentation System (Bio-Plex Workstation from Bio-Rad Laboratories), and each molecule’s concentration was calculated using Bio-Plex Manager Software. The results are expressed in pg/mg protein.
Statistical analysis
Comparisons between experimental groups (PBS- vs. ITC-treated) were performed for each time point (0, 4, 8, and 12 weeks) using a two-tailed Student’s t test with ‘R’ software. Differences between groups were considered statistically significant at p ≤ 0.05 and highly significant at p ≤ 0.01. The immunomodulatory activity of ITC was defined for each molecule.
Results
ITC-treatment induced immunomodulatory effects at the pulmonary level
ITC treatment exerted a significant modulatory effect on 14/28 (50 %) of the immunological molecules evaluated. Three different patterns of the pulmonary levels of these molecules were also observed.
Changes observed from the first evaluation time point until the end of ITC treatment: This pattern was observed for IL-1β, which decreased significantly during ITC treatment compared with PBS treatment. However, the cytokine returned to normal levels upon completion of treatment.
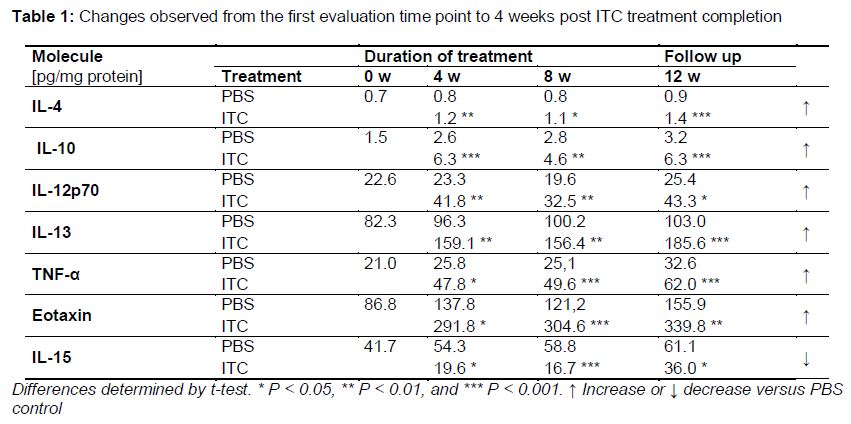
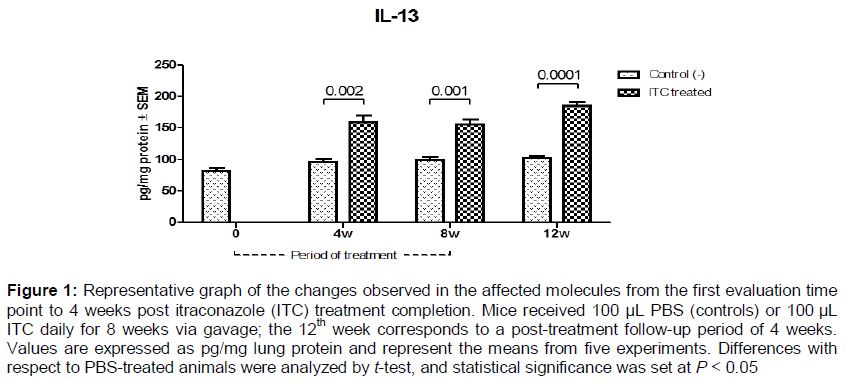
Changes observed from the first evaluation time point until 4 weeks post ITC treatment completion: This pattern was observed for seven molecules, most of which were increased significantly compared with the control group; only IL-15 levels decreasing during ITC treatment (, ).
Changes observed only at the end of ITC therapy
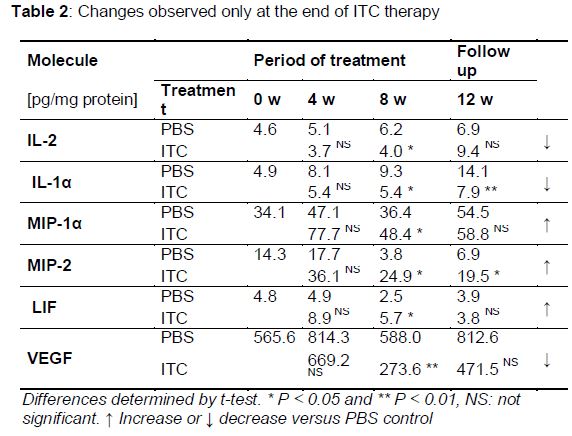
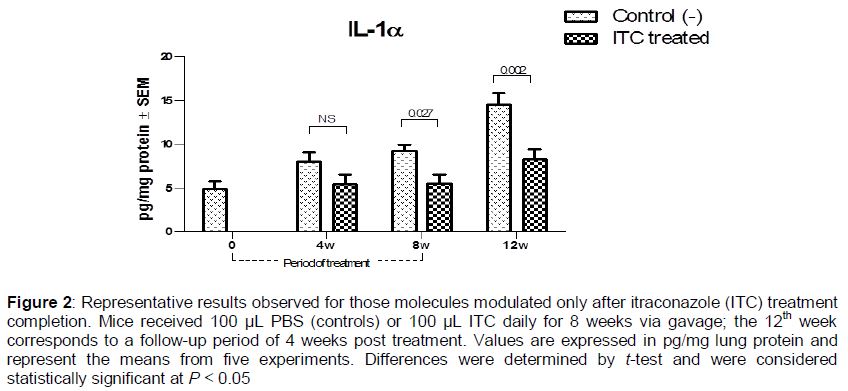
Six molecules did not exhibit changes in their pulmonary levels until after 8 weeks of treatment. Some were increased, while others decreased significantly. The levels of IL-1α and MIP-2 remained altered even at 4 weeks post treatment completion (, ).
Of the 28 molecules evaluated, 14, including cytokines, growth factors, and chemokines (IL-3, IL-5, IL-6, IL-9, IL-17, IL-12p40, G-CSF, GM-CSF, FGF, IFN-γ, MIP-1β KC, MIG, and PDGF), showed no changes induced by ITC treatment at any of the time points evaluated.
Pulmonary histology
Coronal sections of whole lungs stained with H&E were viewed by light microscopy, and the presence of any pulmonary alterations and/or inflammation possibly resulting from ITC treatment was examined. In both PBS-treated (control) and ITC-treated mice, the histology of the lungs was normal and indistinguishable between the groups. The airways remained clear, with the alveoli and alveolar ducts intact and no evidence of damage, alteration, inflammation, or cell migration. Furthermore, we observed no bronchiolar, peribronchiolar, or perivascular inflammation in either group. Representative lung sections from PBS-treated and ITC-treated mice after 8 weeks are shown in .
Discussion
Compared with PBS control group, ITC treatment had a significant modulating effect on the pulmonary levels of 50 % of the molecules evaluated in healthy BALB/c male mice. Some of these molecules changed at the beginning or end of ITC treatment (IL-1α, IL-1β, IL 2, MIP-1α, MIP-2, LIF, and VEGF), while others showed changes throughout the ITC treatment course and even 4 weeks after treatment completion (IL-4, IL-10, IL-12p70, IL-13, IL-15, TNF-α, and eotaxin).
ITC is a synthetic triazole antifungal agent that is widely used because of its broad antimycotic spectrum [1,2,4,9]. Like other triazoles, it is a heterocyclic compound containing a five-membered diunsaturated ring structure composed of three nitrogen atoms and two carbon atoms at non-adjacent positions. This triazole scaffold is extremely versatile and has a broad spectrum of pharmacological activities [15]. Recently, there has been interest in the non-antifungal effects of this group of drugs, because knowledge of these properties could enhance both their clinical usefulness and therapeutic efficacy [3].
Several papers describing the activity of antifungal drugs on the immune system have been published, most based on in vitro studies [7-10]. Regarding the immunomodulatory effects of azoles, the results have been conflicting. Vaughn et al [16] showed that aerosolized ITC given to ICR mice for 12 days did not have inflammatory or modulatory effects in the lungs; they evaluated IL-12 levels in bronchoalveolar lavage fluid and found no change compared with control animals [16]. However, in a review of the immunomodulatory activity of antifungal drugs, Yamaguchi et al [3] suggested that azoles, in general, tend to act as immunosuppressants. Consistent with this, our results showed that levels of IL-1β and IL-1α were decreased significantly in the lungs of ITC-treated mice; these cytokines are considered important molecules in antifungal host defenses and key to induction of the Th1 protective response [6,17]. Additionally, several reports have suggested that the production of Th2 cytokines (IL-4, IL-5, IL-10, and IL-13) is associated with susceptibility to pulmonary fungal infections [18]. Our results showed that pulmonary levels of IL-4, IL-10, and IL-13, considered anti-inflammatory molecules, were higher during and even 4 weeks after the end of ITC treatment in mice.
Although the histopathological analysis showed no change in pulmonary cellular infiltrates, three molecules (MIP-1α, MIP-2, and eotaxin) considered chemokines in polymorphonuclear leukocytes (PMNs) were upregulated after ITC treatment compared with PBS-treated mice. Because PMNs are an important component of the host defense against opportunistic fungi, their suppression could diminish the ability of the host to control this type of infection [19,20].
Sources of these pulmonary cytokines may include several cell types, such as mast cells, epithelial cells, T cells, alveolar macrophages, and even eosinophils, which produce IL-4 and IL-5 [21-22]. We suggest that the pulmonary microenvironment may be defined by the presence of local effector cells and the cytokine milieu that they create, or, as observed in this study, by the use of certain medications that can influence the type of response generated. The precise mechanism(s) by which ITC exerts its effects on the immune system and whether the corresponding changes have clinically significant results remain unclear. One possible mechanism is the ability of ITC to inhibit cytochrome P450 enzyme systems [23]. ITC has been shown to inhibit CYP450 enzyme 5-lipooxigenase dose-dependently, interfering with the synthesis of leukotriene B4 (LTB4), the major product of arachidonic acid metabolism, which has been implicated as a potent mediator of inflammatory processes and immunoregulation [23]. In addition, it has been demonstrated in primary cultures of human hepatocytes that some azolic drugs are antagonists of the human glucocorticoid receptor (GR), which belongs to the family of steroid/thyroid receptors. GR regulates many cellular functions such as homeostasis, cell differentiation, proliferation, and host immune defenses [24]. These properties could explain the antiinflammatory activity of ITC observed in chronic granulomatous disease [25] and allergic bronchopulmonary aspergillosis [26]. Additionally, given the molecular structure of imidazole antimycotics, ITC has been shown to have more potent anti-inflammatory activity than those of ketoconazole, fluconazole, and voriconazole [18]. Recently, Lindstedt et al evaluated the therapeutic effects of a triazole derivative (ST1959) on an autoimmune disease; they found that this triazole decreased human T cell proliferation and inhibited the expression of some cytokines at the transcriptional level, specifically by inhibition of the nuclear localization of nuclear factor of activated T cells [27].
Conclusion
The findings indicate that ITC, at a serum level of 1 μg/mL (therapeutically achievable concen-tration), may exert a marked immunomodulatory effect, accompanied by a slight immunosuppressive tendency at the pulmonary level in healthy male BALB/c mice. These results suggest an alternative, albeit unexplored, mechanism of ITC-mediated immunomodulation. Further studies evaluating more time points and parallel pharmacokinetic analyses are needed in the future to examine this immunomodulatory activity.
Declarations
Acknowledgement
References
Archives


News Updates